The Pleistocene Coyote from the Río Nacaome, Costa Rica
El coyote del Pleistoceno del río Nacaome, Costa Rica
Spencer G. Lucas1* & Guillermo E. Alvarado2
1New Mexico Museum of Natural History and Science, Albuquerque, New Mexico, USA
2Universidad de Costa Rica (UCR), Research Center in Geologial Sciences (CICG by its acronym in Spanish), San José, Costa Rica
*Corresponding author: spencer.lucas@dca.nm.gov
(Recibido: 05/07/2025; aceptado: 12/08/2025)
Revista Geológica de América Central, 73, 2025, doi: https://doi.org/10.15517/ytzr8902
ISSN: 0256-7024
Abstract: In 1997, we identified as coyote (Canis latrans) a dentulous left dentary of a canid from the Upper Pleistocene of the Río Nacaome in northwestern Costa Rica. This identification has been questioned based on misconceptions about features of the dentary that supposedly diagnose C. latrans. We reaffirm the identity of the Río Nacaome canid as C. latrans based on long established features of the lower dentition that are diagnostic of C. latrans. We also refute the idea that certain features of the dentary are diagnostic of coyotes. The Nacaome coyote fossil is the only fossil of a coyote from Central America and establishes their late Pleistocene presence well south of records in southern Mexico.
Keywords: Pleistocene, coyote, Costa Rica, dentary, Canis latrans, C. familiaris.
Resumen: En 1997, identificamos como coyote (Canis latrans) un dentario izquierdo dentuoso de un cánido del Pleistoceno superior del río Nacaome, en el noroeste de Costa Rica. Esta identificación ha sido cuestionada debido a conceptos erróneos sobre las características del dentario que supuestamente diagnostican a C. latrans. Reafirmamos la identidad del cánido del río Nacaome como C. latrans con base en características establecidas desde hace tiempo de la dentición inferior que son diagnósticas de C. latrans. También refutamos la idea de que las características del dentario sean diagnósticas de los coyotes. El fósil de coyote de Nacaome es el único fósil de un coyote de América Central y establece su presencia en el Pleistoceno tardío bastante más al sur de los registros en el sur de México.
Palabras clave: Pleistoceno, coyote, Costa Rica, dentario, Canis latrans, C. familiaris.
Introduction
Coyotes (Canis latrans) are New World canids that now live in diverse habitats from Alaska to Panama (e. g., Bekoff y Gese, 2003; Macdonald y Sillero-Zubiri, 2004; Hody y Kays, 2018). Ancestral coyotes (Canis lepophagus) were widespread in North America by late Pliocene time, and the species C. latrans appeared during the Pleistocene and has an extensive fossil record in the United States and Mexico (Arroyo-Cabrales y Carranza, 2009; Tedford et al., 2009).
Although coyotes now live across much of Central America, their Central American fossil record consists of one dentary from the Upper Pleistocene vertebrate fossil locality on the Río Nacaome in the Nicoya Peninsula of northern Costa Rica (Lucas et al., 1997). Jiménez et al. (2022; also see Gómez et al., 2015) recently questioned identifying this dentary as coyote (Canis latrans), claiming it actually belongs to a domestic dog (C. familiaris). Here, we re-evaluate the taxonomic status of the Río Nacaome dentary to confirm its identification as C. latrans.
Provenance
The coyote dentary is from the Río Nacaome (also called Barra Honda) vertebrate-fossil locality on the Nicoya Peninsula (coordinates 10°8’52.91’’N, -85°19’24.27’’W, Matambú sheet, 10 m a.s.l. ). The dentary is in the collection of the Museo Nacional de Costa Rica in San José, Costa Rica, catalogued as MNCR G24 NC-65 (Fig. 1). Vega (1993) and Valerio (1995) described the geological context of this fossil site. At the site, fossil bones are scattered over an area of 20 m2. They are disarticulated and somewhat aligned, indicating fluvial transport. Valerio (1995) noted that the bones are mineralized and lack collagen suitable for radiocarbon dating. However, such dating may be possible in the future because of recent advances in analytical techniques that have enabled recovery and analysis of micro samples of collagen.
The Nacaome fossil assemblage was recovered in deposits of fluvial origin, particularly from coarse-grained beds with a maximum thickness of 1.2 m consisting of sandstone and shale lithoclasts ≤ 5 cm in diameter with flints up to 15 cm in diameter that are subrounded yellow and red in color. The base of the deposit, on the sedimentary rocks of the Mesozoic-Cenozoic (K-Pg) boundary, is represented by a hard white layer, possibly calcrete. The conglomerate with megafauna is overlain by a similar conglomerate, but with a lower percentage of flint, and the maximum diameter of the lithoclasts is 3-10 cm; it does not contain megafaunal remains. A sandy and clayey soil 1.5 m thick overlies the entire conglomeratic sequence. These lenticular deposits are interpreted as possible point bars, interdigitating with beds of fine sand and floodplain clays. The total thickness does not exceed 10 m, and most of the remains were recovered at a depth of 1–2 m. The alluvial sediments have a primary inclination of 6° to the south-southeast (45°W) (Vega, 1993; Acuña-Mesén and Laurito-Mora, 1996).
The first fossil discovery at the Río Nacaome site was of the jaw and tusk of what has been called the Nacaome or Barra Honda “mastodon,” which was found in a Quaternary fluvial conglomerate, although unfortunately no further data were provided on the sediments. Subsequently, the National Museum carried out explorations and excavations in this locality, mainly between 1990 and 1997, finding a large number of fossil remains, including a gomphothere (Cuvieronious hyodon), horses (Equus sp.), a monkey (Alouatta sp.), semi-aquatic turtle (Rhinoclemmys nicoyana), glyptodont (Glyptotherium sp.), coyote (Canis latrans), a fish dentary (Teleostei ident.), and freshwater bivalve (Unio sp.) (Laurito, 1990; Laurito et al., 1993; Valerio, 1995; Acuña-Mesén y Laurito-Mora, 1996; Lucas et al., 1997). The paleoenvironment of the fossil deposit was possibly a wooded savannah forest with marked seasonality and wetlands or seasonal swamps.
The coyote dentary from the Río Nacaome site is clearly a fossil (Fig. 1), and the geological context and associated fossils indicate it is of late Pleistocene age. It is thus within the age range of fossils of Canis latrans found farther north in the USA and Mexico (Nowak, 1979; Arroyo-Cabrales y Carranza, 2009; Tedford et al., 2009).
Nacaome Dentary: Description
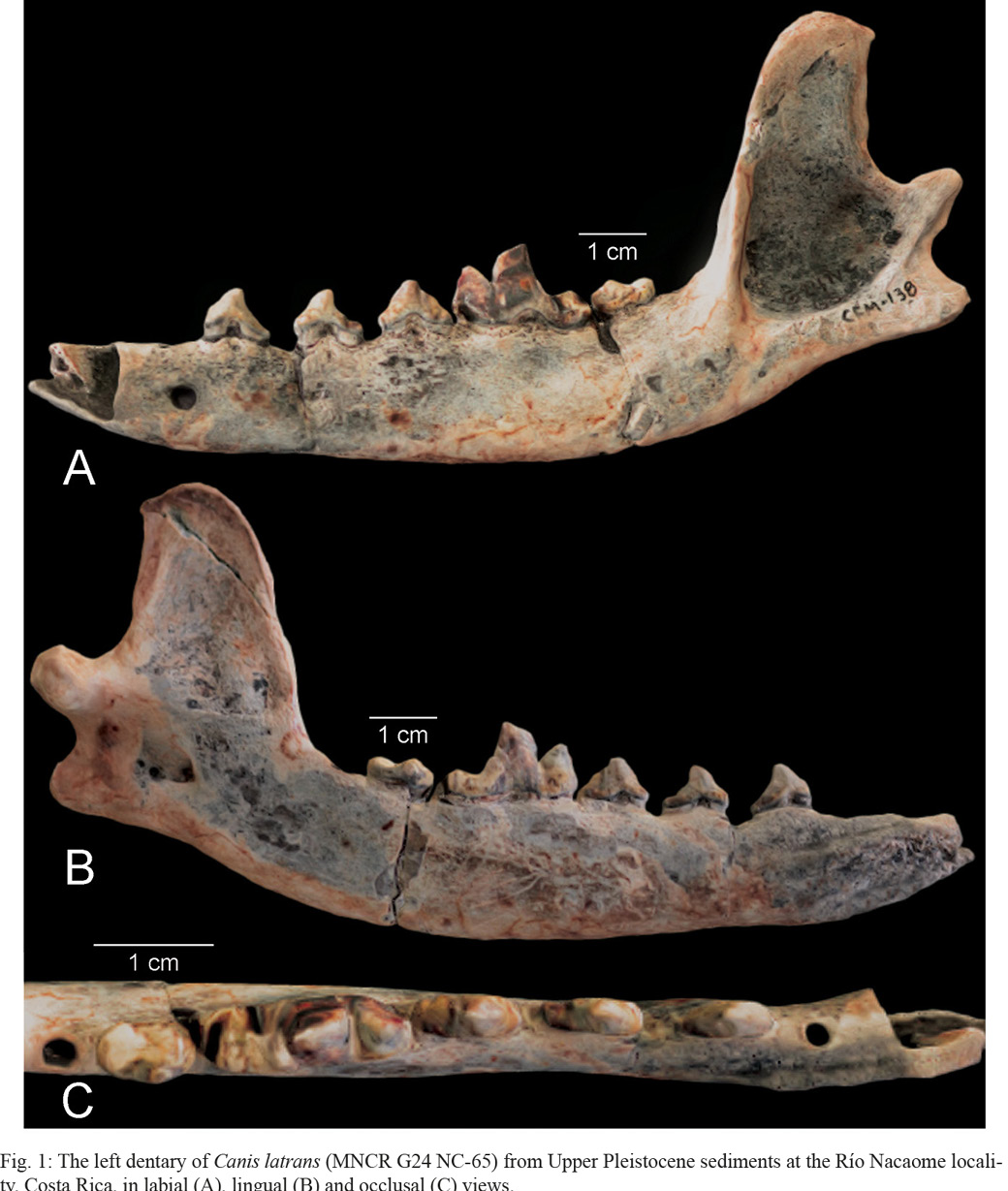
Lucas et al. (1997) presented a brief description, tooth measurements and photographs of the coyote dentary from the Río Nacaome site (dental nomenclature used here follows Tedford et al., 2009, fig. 70). We present here a more detailed description and illustrations of this fossil, which we refer to as the Nacaome dentary. (Fig. 1). The fossil is a left dentary with the alveoli of the c, p1 and m3 and complete p2-m2. It is mineralized, as is clear from the density and coloration of the teeth and dentary.
The anterior edge of the dentary is broken so that the incisors and their alveoli are not preserved, nor is the canine, and the canine alveolus is broken anteriorly (though note that Jimenez et al., 2022, fig. 4B, restored the alveoli). The canine alveolus is large, cylindrical in cross section and directed antero-dorsally. The p2-4 have narrow (trenchant) crowns and are separated from each other by short diastemata. The largest diastema is between p2 and p3. The p2-3 crowns are dominated by a large principal cuspid that is canted slightly posteriad. A single posterior cuspid is present on p2, but p3 has two posterior cuspids. The p4 also has two posterior cuspids and a posterior cingulid to which the posterior cuspids are not connected. The tip of the m1 paraconid is above the level of the entire p4 crown.
The m1 is the large, trenchant carnassial tooth and is followed by a much smaller and lower crowned m2. There is a single, small alveolus for the m3. The m1 protoconid and paraconid are tall and blade like. The anterior face of the paraconid slopes backward. The m1 has a basin-shaped talonid with two prominent cuspids (entoconid and hypoconid), and the talonid is short, about 25% of the length of the entire m1 crown. The m2 has relatively large anterior cuspids (protoconid and metaconid), and well-developed antero-lingual cingulid. The metaconid is smaller than and slightly oblique to the protoconid.
The dentary horizontal ramus is long, slender and slightly convex ventrally beneath p4-m2. A prominent mental foramen is in the lateral aspect of the dentary ventral to the p1 alveolus, and damage to the dentary obscures the possible presence of a second, smaller mental foramen ventral to the p3. The ascending ramus of the dentary is nearly vertical with a coronoid process that is tall with a dorsally curved margin The masseteric fossa is well defined and ovoid to trapezoidal in lateral view. It has well defined anterior and ventral margins. The temporal fossa is much shallower. The superior notch is shallow and open posteriorly. Both the mandibular condyle and angular process are short, robust and project posteriorly. The root of the condyle is an oblique ridge that extends antero-ventrally to the ventral edge of the horizontal ramus.
Nacaome Dentary: Taxonomic Identification
The above description demonstrates that the Nacaome dentary has many features that have long been used by biologists and paleontologists (e. g., Gidley, 1913; Lawrence and Bossert, 1967; Olsen, 1973; Nowak, 1979; Tedford et al., 2009) to distinguish coyotes (Canis latrans) from both domestic dogs (C. familiaris) and wolves (C. lupus):
1. Relatively trenchant (narrow) premolars.
2. Premolars separated by short diastemata, with the p2 isolated from the other premolars by longer diastemata.
3. The posterior part of the p4 is relatively long compared to total tooth length and width.
4. The p4 crown is below the level of the tip of the m1 paraconid.
5. There is a second posterior cuspid behind the main cuspid on the p4.
6. The second posterior cuspid on the p4 is independent of the posterior cingulid.
7. The m1 has an anteriorly directed preparacrista medial to the parastyle.
8. The m1 (carnassial) has a basin-shaped talonid with two prominent cuspids that is approximately of the same width as the trigonid.
9. The length of the m1 talonid is about one third or less of total m1 length.
10. The protoconid and paraconid of m1 are blade-like.
11. The anterior face of the m1 paraconid slopes backward.
12. The lingual cuspids of the m1 talonid and anterior pair of cuspids on the m2 are relatively large.
13. The m1 entoconid is at the postero-lingual corner of the talonid oblique to the hypoconid, and those cuspids are united by cristids.
14. The m1 talonid basin is closed lingually by the entoconulid crest.
15. The m2 has a well developed antero-lingual cingulid that passes posteriorly across the protoconid on the talonid.
16. The m2 metaconid is only slightly oblique to and smaller than (not significantly enlarged over the size of) the protoconid.
These features justify assignment of the Nacaome dentary to Canis latrans.
Not a Coyote?
Jiménez et al. (2022; also see Gómez et al., 2015) argued that the Nacaome dentary is not that of a coyote, but that of a domestic dog. They claimed (we translate their Spanish text to English here) that the specimen “….is assumed to be the southernmost evidence of populations of this canid [C. latrans], being considered an extraordinary and controversial case because it was assumed to be a coyote from the late Pleistocene, very far from its natural habitat” (p. 56). However, coyotes live today in diverse habitats, including in Costa Rica and Panama (e. g., Monge-Nájera y Morera-Brenes, 1986; Bekoff y Gese, 2003; Wilson y Rutledge, 2020; Barboza-Arias y Charão-Marques, 2024; Monroy-Vilchis et al., 2024), so the Nacaome canid was not “very far from its natural habitat” (Jimenez et al., 2022, p. 56). Indeed, coyotes now live in the Nicoya Peninsula (Romero, 2020-2023).
Jiménez et al. (2022, p. 56-58) then justified identifying the Nacaome dentary (referring to it as “CFM-1387”) as a dog, not a coyote, based on these features:
1. “Regarding CFM-1387, its morphology undoubtedly places it as a dog and the presence of the entire dental battery indicates that it was a specimen with hair, an option that refutes the idea that it was a coyote. The certainty of this statement only requires the comparison of this dentary with those of a coyote and a pre-Hispanic dog (Figure 7).12.”
2. “The key element of this statement is the apex of the coronoid process (Figures 5 and 7), which has been considered to be of irrefutable diagnostic value for several decades, and the curved shape of the dentary ramus is also similar to that of the dog (Figure 5). On the other hand, the masseteric fossa, with a smooth surface and, in appearance, shallow, is usual in females. Thus, the morphology tells us, with a fair amount of certainty, that this element belonged to an adult female dog with a body covered with hair.”
3. “Since the dentary of CFM-1387 does not show a curvature as pronounced as that of the pre-Hispanic dog presented, some might think that perhaps it is a hybrid of coyote and dog, but in these cases, the coronoid processes show a very different shape, so that this option has no morphological support (Figure 8).”
4. “If we look at the data in Table 3, the most interesting thing is that the measurements of the Nacaome individual are between 9 and 10% larger than the average observed in coyotes and between 8 and 18% larger than that of pre-Hispanic dogs.* The conclusion is that this dentary belonged to a dog with an elongated face, with a head of about 19 centimeters in length.”
5. “As can be seen, the length of the jaw is where the greatest difference with respect to dogs is observed (around 18%), which justifies its long-faced condition, but in the remaining measurements the difference is 8 to 10%, which means that it was a dog with an elongated but not robust head, that is, with a dolichocephalic tendency. In this sense it undoubtedly has a certain resemblance to coyotes, but the morphological data presented are indisputable in its taxonomic condition, which clearly indicates that it was a dog with an elongated face.”
They noted, “Regarding the morphology of the dentary, as can be seen in figures 2 and 5, we have a mandibular ramus whose lower edge forms an arch and, most importantly, an apex is clearly visible on the posterior edge of the upper end. Finally, the masseteric fossa, in appearance, does not have internal edges.”
However, these statements are inconsistent with nearly 100 years of paleontological and biological studies of coyotes, from Gidley (1913) to Tedford et al. (2009), which do not distinguish coyotes by the morphological features detailed by Jiménez et al. (2022). Thus, works by Gidley (1913), Howard (1949), Jackson (1951), Lawrence y Bossert (1967), Olsen (1973), Kurtén (1974), Nowak (1979), Tedford et al. (2009) and Ayoob (2016) distinguish coyotes by dental characters, not by those of the dentary. The fact is that the dentaries of coyotes are not distinct from those of some other canids, including foxes and many domestic dogs (see especially Olsen, 1973, figs 19-20). Thus, to respond to the points made by Jiménez et al. (2022):
1. Coyotes have a complete lower cheek tooth dentition of p1-m3.
2. The shape of the apex of the dentary coronoid process has never been seen of as of “irrefutable diagnostic value” in identifying coyotes (see, especially Gidley, 1913 and Tedford et al., 2009). Jiménez et al. (2022) cite Valadez et al. (2010) as the source of this claim, but a careful reading of Valadez et al. (2010) reveals no discussion or analysis of the use of the dentary coronoid process to distinguish coyotes from other canids.
3. Similarly, curvature of the dentary horizontal ramus has never been used to diagnose coyotes from dogs.
4. The claim that measurements of the Nacaome jaw has teeth that are 9-10% larger than those of coyotes is based on a very limited sample (Jiménez et al., 2022, table 3). Tedford et al. (2009) presented dental measurements of much larger samples of fossil and extant coyotes, and the Nacaome dentary has teeth with dental measurements within the ranges or extremely close to the ranges of those measurements (Table 1). Furthermore, Pleistocene coyotes are generally larger than Recent coyotes (Meachen y Samuels, 2012; Meachen et al., 2014; Llano-Enderle y Ruiz-Ramoni, 2021).
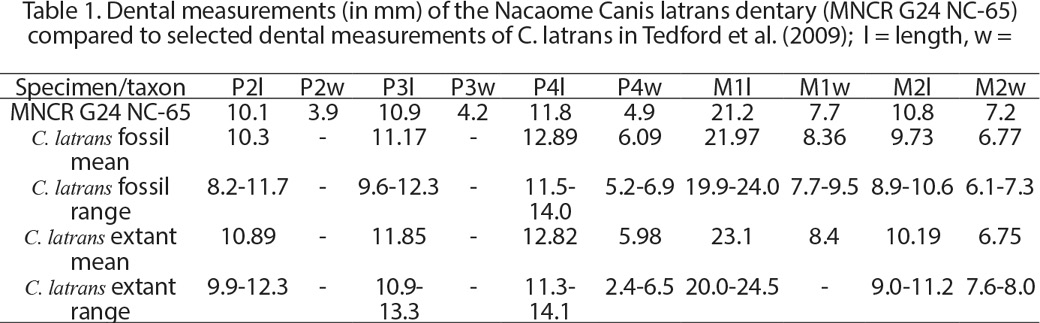
5. The claim that the length of the mandible is a major difference between dogs and coyotes is also questionable. Coyotes are dolicochepalic, and this is seen in their relatively long and slender dentaries. Jiménez et al. note that the Nacaome dentary is relatively long and slender but dismiss it as taxonomically insignificant.
6. Their statement that the Nacaome dentary has a masseteric fossa lacking “internal edges” is not clear to us. But, note that the masseteric fossa of the Nacaome dentary does not differ from the fossa on coyote dentaries (Figs 2-3).
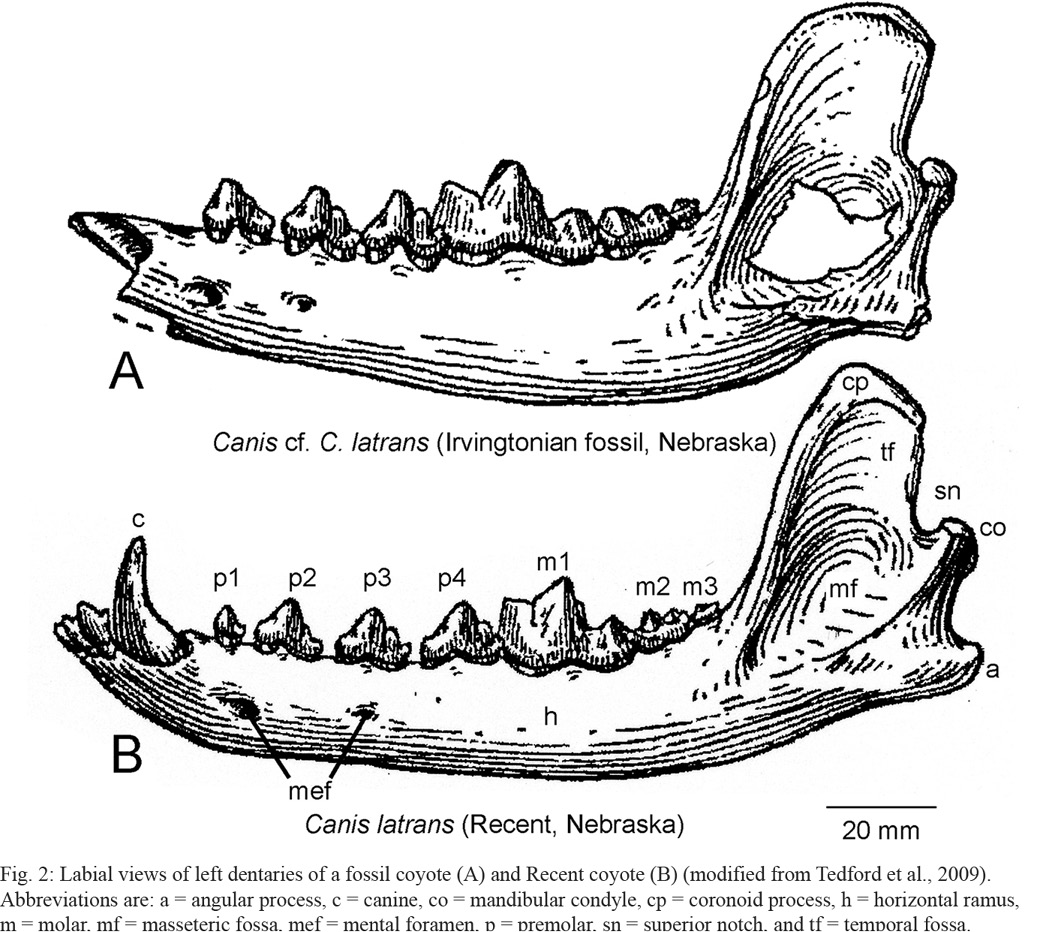
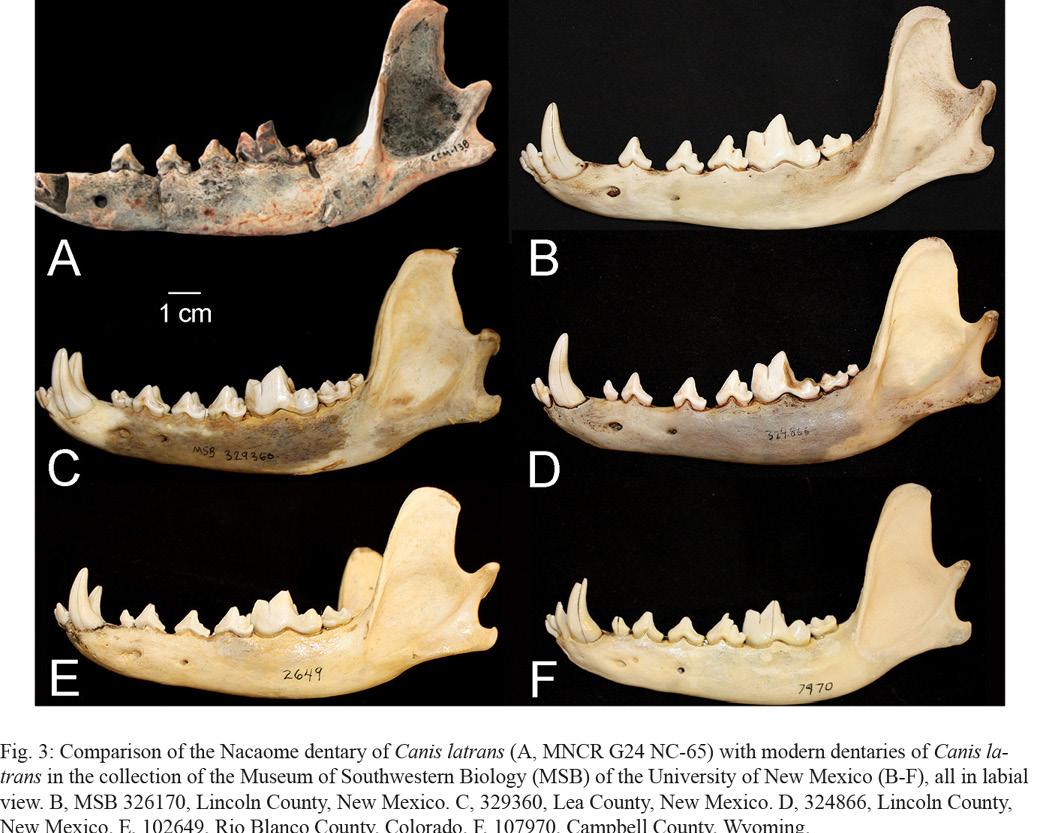
Dentaries of coyotes, fossil and extant, vary slightly in some of the features that Jiménez et al. (2022) discuss (Figs. 2-3). Thus, the overall shape—length, height and dorsal margin—of the coronoid process varies from long and rounded dorsally (Figs 2A, 3E) to much shorter and pointed dorsally, as in the Nacaome dentary (Figs. 2B, 3B, F). Also, the degree of ventral curvature of the horizontal ramus varies somewhat. Indeed, our comparison of the Nacaome dentary to a large sample of extant coyote dentaries from the USA in the MSB collection (Fig. 3) reveals a range of variation that includes extant coyote dentaries virtually identical in all morphological features to the Nacaome dentary (compare Figs 3A and B).
We conclude that Jiménez et al. (2022) present an erroneous analysis of the morphology of the dentary and lower dentition to distinguish coyotes from dogs. Features of the dentary that they claim distinguish coyotes (Canis latrans) from dogs (C. familiaris) are an invention of their work, not observations supported by a century of careful research.
Conclusion
Coyotes are canids native to the Americas with a well established Pleistocene fossil record in the USA and Mexico. The Pleistocene occurrence of a coyote in Costa Rica is not surprising, and if the Central American vertebrate fossil record did not suffer from a taphonomic megabias against fossils of small mammals, we believe there would be more coyote fossils from the Central American Pleistocene (Lucas et al., 2022). We should also note that the oldest New World dogs (C. familiaris) are no older than about 10 kyr (Ní Leathlobhair et al, 2018; Perri et al., 2019, 2021), and the late Pleistocene Nacaome dentary is older, and very likely significantly older, than that.
Coyotes now range from Alaska to Panama, much of that range having been extended during the Holocene, and still ongoing (e.g., Hody y Kays, 2018). Late Pleistocene records of coyotes in southern Mexico and Costa Rica reflect their first arrival in the New World tropics.
Acknowledgments
We thank Ana Lucía Valerio at the Museo Nacional de Costa Rica, San José, Costa Rica, and Jonathan Dunnum of the Museum of Southwestern Biology of the University of New Mexico, Albuquerque, New Mexico, USA, for access to collections. Gary Morgan provided helpful comments on an earlier version of the manuscript, and review comments by Richard S. White improved the content and clarity of the manuscript.
References
Acuña-Mesén, R. A., & Laurito-Mora, C. A. (1996). Nueva especie de Rhinoclemmys Fitzinger, 1836 (Chelonii, Cryptodira) del Cenozoico Tardío de Costa Rica. Ameghiniana, 33(3), 271-278.
Arroyo-Cabrales, J., & Carranza, O. (2009). Los canidos prehistóricos de México antes de la llegada del perro. Archaeobios, 1, 34–45.
Ayoob, S. (2016). Comparative Morphology of the Mandible Within Canidae. (Masters thesis). Southern Illinois University.
Barboza-Arias, L. M., & Charão-Marques, F. (2024). Del “fuera de lugar” a los territories excéntricos: coyotes y relacionamientos multiespecie en San Gerardo de Oreamuno, Costa Rica. Horizontes Antropológicos, 70, e700409. https://doi.org/10.1590/1806-9983e700409
Bekoff, M., & Gese, E. M. (2003). Coyote. In G. A. Feldhamer, B. C. Thompson & J. A. Chapman (eds.), Wild Mammals of North America: Biology, Management, and Conservation (pp. 467-481). Johns Hopkins University Press. https://doi.org/10.56021/9780801874161
Gidley, J.W. (1913). Preliminary report on a recently discovered Pleistocene cave deposit near Cumberland, Maryland. Proceedings of the United States National Museum, 46, 93–102. https://doi.org/10.5479/si.00963801.46-2014.93
Gómez M, Espinosa A, Rodríguez B, Valadez R, Ortega M, Viniegra F., & Olmos K. (2015). ¿El mejor amigo del hombre? Como vivieron cánidos prehiapánicos y modernos encontrados en el Barrio Oaxaqueño, Teotihuacan. AMMVEPE, 26(1), 5-21.
Hody, J. W., y Kays, R. (2018). Mapping the expansion of coyotes (Canis latrans) across North and Central America. Zookeys, 759, 81-97. https://doi.org/10.3897/zookeys.759.15149
Howard, W. E. (1949). A means to distinguish skulls of coyotes and domestic dogs. Journal of Mammalogy, 30, 169-171. https://doi.org/10.2307/1375266
Jackson, H. H. T. (1951). Classification of the races of the coyote. Part II. In. S. P. Young, and H. H. T. Jackson (eds.), The Clever Coyote (pp. 227-441). Wildlife Management Institute.
Jiménez, G. V., Báez-Sirias, M., & Valadez-Azua, R. (2022). Cánidos de la región de Gran Nicoya, Costa Rica. Parte uno: Perra pleistocénica (Canis lupus familiaris) en el sitio de Nacaome y coyote prehispánico (Canis latrans) de Jícaro, Guanacaste. AMMVEPE, 33(2), 53-64.
Kurtén, B. (1974). A history of the coyote-like dogs (Canidae; Mammalia). Acta Zoologica Fennica, 140, 1–37.
Laurito, C. A. (1990). Estudio de un ejemplar de Cuvieronius hyodon hallado en el lecho del Rio Nacaome, Guanacaste, Costa Rica. Revista Geologica de America Central, 11, 41-50. https://archivo.revistas.ucr.ac.cr/index.php/geologica/article/view/13009/12287
Laurito, C. A., W. Valerio, &Vega, E. (1993). Nuevos hallazgos paleovertebradológicos en la Peninsula de Nicoya: implicaciones paleoambientales y culturales de la fauna de Nacaome. Revista Geológica de América Central, 16:113-115. https://archivo.revistas.ucr.ac.cr/index.php/geologica/article/view/13280/12549
Lawrence, B., & Bossert, W. H. (1967). Multiple character analysis of Canis lupus, latrans, and familiaris, with a discussion of the relationships of Canis niger. American Zoologist, 7, 223-232. https://doi.org/10.1093/icb/7.2.223
Ní Leathlobhair, M., Perri, A. R., Irving-Pease, E. K., Witt, K. E., Linderholm, A., Haile, J., Lebrasseur, O., Ameen, C., Blick, J., Boyko, A. R., Brace, S., Cortes, Y. N., Crockford, S. J., Devault, A., Dimopoulos, E. A., Eldridge, M., Enk, J., Gopalakrishnan, S., Gori, K., … Frantz, L. A. F. (2018). The evolutionary history of dogs in the Americas. Science, 361(6397), 81-85. https://doi.org/10.1126/science.aao4776
Llano-Enderle, R. A. & Ruiz-Ramoni, D. (2021). Variation in dental size between Pleistocene and living coyotes ( Canis latrans Say, 1823) from Mexico. Journal of South American Earth Sciences, 111, 103500. https://doi.org/10.1016/j.jsames.2021.103500
Lucas, S. G., Alvarado, G. E., & Vega, E. (1997). The Pleistocene mammals of Costa Rica. Journal of Vertebrate Paleontology, 17, 413-427. https://doi.org/10.1080/02724634.1997.10010985
Lucas, S. G., Romero, J. E., Vásquez, O. J., & Alvarado, G. E. (2022). The fossil vertebrates of Guatemala. Revista Geológica de América Central, 66, 1-32. https://doi.org/10.15517/rgac.v66i0.48590
Macdonald, D. W., & Sillero-Zubiri, C. (2004). The biology and conservation of wild canids. Oxford University Press Inc. https://doi.org/10.1093/acprof:oso/9780198515562.001.0001
Meachen, J. A., & Samuels, J. X. (2012). Evolution in coyotes ( Canis latrans) in response to the megafaunal extinctions. PNAS, 109, 4191-4196. https://doi.org/10.1073/pnas.1113788109
Meachen, J. E., Janowicz, A. C., Avery, J. E., & Sadleir, R. W. (2014). Ecological changes in coyotes (Canis latrans) in reponse to the Ice Age megafaunal extinctions. PloS ONE, 9(12), e116041. https://doi.org/10.1371/journal.pone.0116041
Monge-Nájera, J., y Morera-Brenes, B. (1986). La dispersion del coyote ( Canis latrans) y la evidencia de los antiguos cronistas. Brenesia, 25-26, 251-260.
Monroy-Vilchis, O., González-Maya, J. F., Balbuena-Serrano, A., Elvir, F., Zarco-González, M. M., & Rodríguez-Soto, C. (2024). Coyote ( Canis latrans) in South America: potential routes of colonization. Integrative Zoology, 15, 471-481. https://doi.org/10.1111/1749-4877.12446
Nowak, R. M. (1979). North American Quaternary Canis. Monograph of the Museum of Natural History, University of Kansas, 6, 1-154. https://doi.org/10.5962/bhl.title.4072
Olsen, S. J. (1973). Mammal remains from archaeological sites. Part I. Southeastern and southwestern United States. Papers of the Peabody Museum of Archaeology and Ethnology, 56, 1-162.
Perri, A., Widga, C., Lawler, D., Martin, T., Loebel, T., Farnsworth, K., Kohn, L., & Buenger, N. (2019). New evidence of the earliest domestic dogs in the Americas. American Antiquity, 84, 68-87. https://doi.org/10.1017/aaq.2018.74
Perri, A. R., Feuerborn, T. R., Frantz, L. A. F., Larson, G., Malhi, R. S., Meltzer, D. J., & Witt, K. E. (2021). Dog domestication and the dual dispersal of people and dogs into the America. PNAS, 118(6), e2010083118. https://doi.org/10.1073/pnas.2010083118
Romero, Y. A. (2020-2023). Áreas de actividad y dieta del Canis latrans en el contexto etnoecológico de los bosques tropical seco-transición húmedo, bosque muy húmedo, premontano, montano y subalpino en Costa Rica. (Informe final). Sinac. https://www.researchgate.net/publication/377491458_Informe_final_Areas_de_actividad_y_dieta_del_Canis_latrans_en_el_contexto_etnoecologico_de_los_bosques_tropical_seco-transicion_humedo_bosque_muy_humedo_premontano_montano_y_subalpino_en_Costa_Rica
Valadez, R., Götz, Ch., & Mendoza, V. (2010). El perro pelón, su origen, su historia. Instituto de Investigaciones Antropológicas de la Universidad Nacional Autónoma de México, Universidad Autónoma de Yucatán.
Valerio, W. L. (1995). Marcas y hundimientos en huesos de fauna Pleistocénica de Nacaome, Guanacaste. Vínculos, 21, 79-98.
Vega, E. (1993). Geología y aspectos paleontológicos de la excavación realizada en el Río Nacaome. Museo Nacional de Costa Rica Departamento de Historia Natural Sección de Geología.
Wilson, P. J., & Rutledge, L. Y. (2021). Considering Pleistocene North American wolves and coyotes in the eastern Canis origin story. Ecology and Evolution, 11(13), 9137-9147. https://doi.org/10.1002/ece3.7757