Potential distribution and fundamental niche of Moniliophthora spp in cocoa of America and Africa
DOI:
https://doi.org/10.15517/am.v30i3.35038Keywords:
Biogeography, Theobroma cacao, microbial ecology, pathogenic fungi, host pathogen relationsAbstract
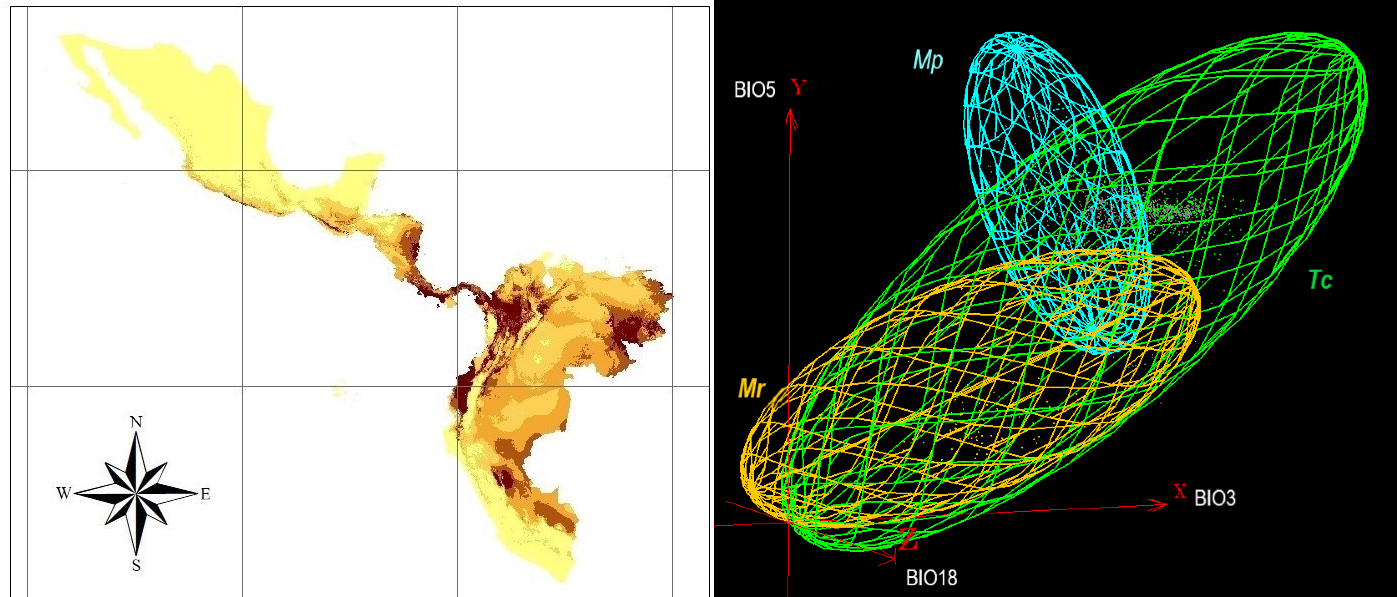
Introduction. Moniliophthora spp. species are endemic from South America and infect America’s cocoa crops. Objective. The objective of the study was to analyze the potential distribution and the fundamental niche of Moniliophthora roreri (Mr) and M. perniciosa (Mp) in America and Africa. Materials and methods. The occurrence records were obtained from a bibliographic review. Bioclimatic variables were used as predictors in the maximum entropy niche model (MaxEnt) and for its visualization in Niche Analyst 3.0. Results. The areas where Mr was most likely to occur were Esmeraldas and Los Ríos, Ecuador, coastal zones of Costa Rica, Santander and Antioquia of Colombia, and for Mp the States of Bahia and Acré of Brazil. The variables that most influenced the distribution of Mr are those associated with precipitation. Temperature-related conditions were more important in the Mp model. The risk of Mr invasion was identified in the Acre and Pará regions, Brazil. The transfer of the models evidenced the potential of Mr and Mp invasion in cocoa crops in Africa. The fundamental niche of pathogens and cocoa showed average similarity values, while the niche of fungal species showed a low similarity. Low pathogen niche similarity and differences in bioclimatic variables affecting pathogens distribution were related to a process of niche partitioning. Conclusion. Geographical regions with a greater potential distribution of pathogens and bioclimatic characteristics that would favor their probability of presence in America’s cocoa crops were identified. It is necessary to regulate the transport of contaminated plant material and equipment between regions to avoid the introduction of pathogens in healthy cocoa crops.
Downloads
References
Adler, P.B., A. Fajardo, A.R. Kleinhesselink, and N.J. Kraft. 2013. Trait-based tests of coexistence mechanisms. Ecol Lett. 16:1294-1306. doi:10.1111/ele.12157
Agrios, G.N. 2005. Plant pathology. 5th ed. University of Florida, FL, USA.
Aime, M.C., and W. Phillips-Mora. 2005. The causal agents of witches’ broom and frosty pod rot of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycologia 97:1012-1022.
Ali, S.S., J. Shao, M.D. Strem, W. Phillips-Mora, D. Zhang, L.W. Meinhardt, and B.A. Bailey. 2015. Combination of RNAseq and SNP nanofluidic array reveals the center of genetic diversity of cacao pathogen Moniliophthora roreri in the upper Magdalena Valley of Colombia and its clonality. Front. Microbiol. 6:850. doi:10.3389/fmicb.2015.00850
Andebrhan, T., A. Figueira, M.M. Yamada, and D.B. Furtek. 1999. Molecular fingerprinting suggests two primary outbreaks of witches’ broom disease (Crinipellis perniciosa) of Theobroma cacao in Bahia, Brazil. Eur. J. Plant Pathol. 105:167-175. doi:10.1023/A:1008716000479
Artero, A.S., J.Q. Silva, P.S. Albuquerque, E.A. Bressan, G.A. Leal, A.M. Sebbenn, G.W. Griffith, and A. Figueira. 2017. Spatial genetic structure and dispersal of the cacao pathogen Moniliophthora perniciosa in the Brazilian Amazon. Plant Pathol. 66:912-923. doi:10.1111/ppa.12644
Barbosa, C.S., R.R. Fonseca, T.M. Batista, M.A. Barreto, C.S. Argolo, M.D. Carvalho, D.O. Amaral, E.M. Silva, E. Arévalo-Gardini, K.S. Hidalgo, G.R. Franco, C.P. Pirovani, F. Micheli, and K.P. Gramacho. 2018. Genome sequence and effectorome of Moniliophthora perniciosa and Moniliophthora roreri subpopulations. BMC Genomics 19:509. doi:10.1186/s12864-018-4875-7
Bartley, G.D. 2005. The genetic diversity of cacao and its utilization. CABI Publishing, Wallingford, GBR.
Bogosian, V., E.C. Hellgren, M.W. Sears, and R.W. Moody. 2012. High-resolution niche models via a correlative approach: Comparing and combining correlative and process-based information. Ecol. Model. 237-238:63-73. doi:10.1016/j.ecolmodel.2012.04.017
Brown, J.L. 2014. SDMtoolbox: a python-based GIS toolkit for landscape genetic, biogeographic, and species distribution model analyses. Methods Ecol Evol. 5:694-700. doi:10.1111/2041-210X.12200
CABI. 2016. Global module. 7nd ed. CABI, Oxfordshire, GRB.
Campuzano, H. 1980. La moniliasis del cacao. Cacaotero Colomb. 13:21-24.
Clement, C.R., M. de-Cristo-Araújo, G.C. d’Eeckenbrugge, A. Alves-Pereira, and D. Picanco-Rodrigues. 2010. Origin and domestication of native Amazonian crops. Diversity 2:72-106. doi:10.3390/d2010072
Cornell, H. 2012. Niche overlap. In: A. Hastings, and L. Gross, editors, Encyclopedia of theoretical ecology (No. 4). University of California Press, CA, USA. p. 489-497.
Cubillos, G. 2017. Frosty pod rot, disease that affects the cocoa (Theobroma cacao) crops in Colombia. Crop. Prot. 96:77-82. doi:10.1016/j.cropro.2017.01.011
Enríquez, G.A., and C.A. Suárez. 1978. Monilia disease of cocoa in Costa Rica. Turrialba 28:339-340.
Escobar, L.E., and M.E. Craft. 2016. Advances and limitations of disease biogeography using ecological niche modeling. Front. Microbiol. 7:1174. doi:10.3389/fmicb.2016.01174
Evans, H.C. 1986. A reassessment of Moniliophthora (Monilia) pod rot of cocoa. Cocoa Growers’ Bull. 37:4-43.
Evans, H.C. 2007. Cacao diseases - the trilogy revisited. Phytopathology 97:1640-1643. doi:10.1094/PHYTO-97-12-1640
Evans, H.C. 2016a. Witches’ Broom disease (Moniliophthora perniciosa): History and biology. In: B. Bayley et al., editors, Cacao diseases: A history of old enemies and new encounters. Springer International Publishing, Basel, CHE. p. 137-177. doi:10.1007/978-3-319-24789-2_5
Evans, H.C. 2016b. Frosty pod rot (Moniliophthora roreri). In: B. Bayley et al., editors, Diseases: A history of old enemies and new encounters. Springer International Publishing, Basel, CHE. p. 63-96. doi:10.1007/978-3-319-24789-2_5
Evans, H.C., J.L. Bezerra, and R.W. Barreto. 2013. Of mushrooms and chocolate trees: aetiology and phylogeny of witches’ broom and frosty pod diseases of cacao. Plant Pathol. 62:728-740. doi:10.1111/ppa.12010
Evans, H.C., D.F. Edwards and M. Rodríguez. 1977. Research on cocoa diseases in Ecuador: Past and present. PANS 23:68-80. doi:10.1080/09670877709412399
Evans, H.C., K.A. Holmes, W. Phillips-Mora, and M.J. Wilkinson, M.J. 2002. What’s in a name, Crinipellis, the final resting place for the frosty pod rot pathogen of cocoa? Mycologist 16:148-152. doi:10.1017/S0269-915X(02)00409-3
Fick, S.E., and R.J. Hijmans. 2017. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017:5086. doi:10.1002/joc.5086
GBIF. 2018. Occurrence download. GBIF, DNK. doi:10.15468/dl.jka55n
González, L.C. 1981. Efecto de las fuentes de inóculo sobre las posibilidades de combate de la moniliasis del cacao. Acta de la Primera Jornada de Investigación. Universidad de Costa Rica, San José, CRI.
Gramacho, K.P., L.E.M. Newman, F.S. da-Silva, U.V. Lopes, J.L. Pires, and L. Pereira. 2016. Pathogenic variability of Moniliophthora perniciosa in three agroecological zones of the cacao region of Bahia, Brazil. Crop Breed. Appl. Biotechnol. 16:7-13. doi:10.1590/1984-70332016v16n1a2
Griffith, G.W., J. Nicholson, A. Nenninger, R.N. Birch, and J.N Hedger. 2003. Witches’ brooms and frosty pods: two major pathogens of cacao. New Zeal. J. Bot. 41:423-35. doi:10.1080/0028825X.2003.9512860
Hutchinson, G.E. 1957. Concluding remarks. Cold Spr. Harb. Symp. Quant. Biol. 22:415-442. doi:10.1101/SQB.1957.022.01.039
Jaimes, Y., and F. Aranzazu. 2010. Manejo de las enfermedades del cacao (Theobroma cacao L.) en Colombia, con énfasis en monilia (Moniliophthora roreri). Corpoica, COL.
Kumar, S., L.G. Neven, and W.L. Yee. 2014. Evaluating correlative and mechanistic niche models for assessing the risk of pest establishment. Ecosphere 5(7):1-23. doi:10.1890/ES14-00050.1
Leal, G.A., P.S. Albuquerque, and A. Figueira. 2007. Genes differentially expressed in Theobroma cacao associated with resistance to witches’ broom disease caused by Crinipellis perniciosa. Mol. Plant Pathol. 8:279-292. doi:10.1111/j.1364-3703.2007.00393.x
Leandro, M.E. 2011. Efecto de los factores macro y microclimáticas y las características productivas del cacao sobre la epidemiología de la moniliasis. Tesis M.Sc., CATIE, Turrialba, CRI.
Leandro-Muñoz, M.E., P. Tixier, A. Germon, V. Rakotobe, W. Phillips-Mora, S. Maximova, and A. Jacques. 2017. Effects of microclimatic variables on the symptoms and signs onset of Moniliophthora roreri, causal agent of Moniliophthora pod rot in cacao. PLoS One. 12(10):e0184638. doi:10.1371/journal.pone.0184638
Maddison, A.C., G. Macías, C. Moreira, R. Arias, and R. Neira. 1995. Cocoa production in Ecuador in relation to dryseason escape from pod rot caused by Crinipellis perniciosa and Moniliophthora roreri. Plant Pathol. 44:982-98. doi:10.1111/j.1365-3059.1995.tb02657.x
Meinhardt, L.W., J. Rincones, B.A. Bailey, M.C. Aime, G.W. Griffith, D. Zhang, and G.A.G. Pereira. 2008. Moniliophthora perniciosa, the causal agent of witches’ broom disease of cacao: what’s new from this old foe? Mol. Plant Pathol. 9:577-588. doi:10.1111/j.1364-3703.2008.00496.x
Motamayor, J.C., P. Lachenaud, J.W. da-Silva-e-Mota, R. Loor, D.N. Kuhn, J.S. Brown, and R.J. Schnell. 2008. Geographic and genetic population differentiation of the Amazonian chocolate tree (Theobroma cacao L). PLoS One. 3(10):e3311. doi:10.1371/journal.pone.0003311
Orellana, R. 1956. Occurrence of Monilia pod rot and other cacao diseases in eastern Panama. FAO Plant Protect. Bull. 4:168-169.
Osorio-Almanza, L., O. Burbano-Figueroa, A.M. Arcila, M.A. Vásquez, F. Carrascal-Pérez, y J. Romero. 2017. Distribución espacial del riesgo potencial de marchitamiento del aguacate causado por Phytophthora cinnamomi en la subregión de Montes de María, Colombia. Rev. Colomb. Cienc. Hortic.11:273-285. doi:10.17584/rcch.2017v11i2.7329.
Osorio-Solano, C., C.A. Orozco-Castaño, G.A. López-Gartner, y F.A. Rivera-Páez. 2012. Variabilidad genética de Moniliophthora perniciosa (Stahel) Aime y Phillips-Mora, comb. nov. (Agaricales - Marasmiaceae) en variedades de cacao (Theobroma cacao L.). Acta Agron. 61:93-101.
Patrocínio, N.B., P.C. Ceresini, L.S. Gomes, M.V. Resende, E.G. Mizubuti, and K.P. Gramacho. 2017. Population structure and migration of the witches’ broom pathogen Moniliophthora perniciosa from cacao and cultivated and wild solanaceous hosts in southeastern Brazil. Plant Pathol. 66:900-911. doi:10.1111/ppa.12636
Phillips, S.J., P.R. Anderson, and R.E. Schapire. 2006. Maximum entropy modeling of species geographic distributions. Ecol Model. 190:231-259. doi:10.1016/j.ecolmodel.2005.03.026
Phillips, S.J., and M. Dudík. 2008. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161-175. doi:10.1111/j.0906-7590.2008.5203.x
Phillips-Mora, W. 1986. Evaluación de la resistencia de cultivares de cacao (Theobroma cacao L.) a Moniliophthora roreri (Cif. y Par.). Tesis M.Sc., Universidad de Costa Rica, San José, CRI.
Phillips-Mora, W., M.C. Aime, and M.J. Wilkinson. 2007. Biodiversity and biogeography of the cacao (Theobroma cacao) pathogen Moniliophthora roreri in tropical America. Plant Pathol. 56:911-922. doi:10.1111/j.1365-3059.2007.01646.x
Phillips-Mora, W., F. Baqueros, R.L. Melnick, and B.A. Bailey. 2015. First report of frosty pod rot caused by Moniliophthora roreri on cacao in Bolivia. New Dis. Rep. 31:29. doi:10.5197/j.2044-0588.2015.031.029
Phillips-Mora, W., J. Cawich, W. Garnett, and M.C. Aime. 2006a. First report of frosty pod rot (moniliasis disease) caused by Moniliophthora roreri on cacao in Belize. Plant Pathol. 55:584-584. doi:10.1111/j.1365-3059.2006.01378.x
Phillips-Mora, W., A. Coutiño, C.F. Ortiz, A.P. López, J. Hernández, and M.C. Aime. 2006b. First report of Moniliophthora roreri causing frosty pod rot (moniliasis disease) of cocoa in Mexico. Plant Pathol. 55:584-584. doi:10.1111/j.1365-3059.2006.01418.x
Porras, V.H., y L. González. 1982. Capacidad de liberación de mazorcas enfermas dejadas en el árbol de cacao. Actas de la V Reunión Anual de Agronomía. Colegio de Ingenieros Agrónomos, San José, CRI.
Purdy, L.H., and R.A. Schmidt. 1996. Status of cacao witches broom: biology, epidemiology, and management. Annu. Rev. Phytopathol. 34:573-94. doi:10.1146/annurev.phyto.34.1.573
Qiao, H., A. Peterson, C. Townsend, P. Lindsay, J. Soberón, L. Ji, and L.E. Escobar. 2016. NicheA: creating virtual species and ecological niches in multivariate environmental scenarios. Ecography 39:805-813. doi:10.1111/ecog.01961
Scarpari, L. M., L.W. Meinhardt, P. Mazzafera, A.W.V. Pomella, M.A. Schiavinato, J.C.M Cascardo, and G.A.G. Pereira. 2005. Biochemical changes during the development of witches’ broom: the most important disease of cocoa in Brazil caused by Crinipellis perniciosa. J. Exp. Bot. 56:865-877. doi:10.1093/jxb/eri079
Schmidt, R.A., S.A. Rudgard, A.C. Maddison, and T. Andebrhan. 1993. Comparative epidemiology of the witches’ broom pathosystem. In: S.A. Rudgard et al., editors, Disease management in cocoa. Springer, Dordrecht, NLD. p. 131-156. doi:10.1007/978-94-011-2126-2
Shaw, M.W., and A.E. Vandenbon. 2007. A qualitative host-pathogen interaction in the Theobroma cacao-Moniliophthora perniciosa pathosystem. Plant Pathol. 56:277-285. doi:10.1111/j.1365-3059.2006.01549.x
Silva, P. 1987. Cacau e lagarto e vassoura-de-bruxa: registros feitos por Alexandre Rodrigues Ferreira em 1785 e 1787 na Amazonia. Bol. Tecnol. CEPEC 146:3-21.
Soberón, J. 2007. Grinnellian and Eltonian niches and geographic distributions of species. Ecol. Lett. 10:1115-1123. doi:10.1111/j.1461-0248.2007.01107.x
Soberón, J., and A.T. Peterson. 2005. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodiver. Inform. 2:1-10. doi:10.17161/bi.v2i0.4
Suárez-Contreras, L.Y. 2016. Identificación molecular de aislamientos de Moniliophthora roreri en huertos de cacao de Norte de Santander, Colombia. Acta Agron. 65(1):51-57. doi:10.15446/acag.v65n1.47994
Thomas, E., M. van-Zonneveld, J. Loo, T. Hodgkin, G. Galluzzi G, and J. van-Etten. 2012. Present spatial diversity patterns of Theobroma cacao L. in the Neotropics reflect genetic differentiation in pleistocene refugia followed by human-influenced dispersal. PLoS One. 7(10):e47676. doi:10.1371/journal.pone.0047676
Torres-de-la-Cruz, M., C.F. Ortiz-García, D. Téliz-Ortiz, A. Mora- Aguilera, and C. Nava-Díaz. 2011. Temporal progress and integrated management of frosty pod rot (Moniliophthora roreri) of cocoa in Tabasco, Mexico. J. Plant Pathol. 93:31-36. doi:10.4454/jpp.v93i1.270
Villamil, J., L. Sierra, Y. Olarte, A. Mosquera, J. Fajardo, E. Pinzon, E. Mosquera, y J. Martínez. 2015. Integración de prácticas culturales y control biológico para el manejo de Moniliophthora roreri CIF & PAR. Rev. Cienc. Agr. 32(2):13-25. doi:10.22267/rcia.153202.9
Violle, C., D.R. Nemergut, Z. Pu, and L. Jiang. 2011. Phylogenetic limiting similarity and competitive exclusion. Ecol. Lett. 14:782-787. doi:10.1111/j.1461 0248.2011.01644.x
Wiens, J.J., D.D. Ackerly, A.P. Allen, B.L. Anacker, L.B. Buckley, H.V. Cornell, E.I. Damschen, T. Jonathan-Davies, J. Grytnes, S.P. Harrison, B.A. Hawkins, R.D. Holt, C.M. McCain, and P.R. Stephens. 2010. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 13:1310-1324. doi:10.1111/j.1461-0248.2010.01515.x
Zhang, D., and L. Motilal. 2016. Origin, dispersal, and current global distribution of cacao genetic diversity. In: B. Bayley et al., editors, Diseases: A history of old enemies and new encounters. Springer International Publishing, Basel, CHE. p. 3-31. doi:10.1007/978-3-319-24789-2_1
Downloads
Additional Files
Published
How to Cite
Issue
Section
License
1. Proposed policy for open access journals
Authors who publish in this journal accept the following conditions:
a. Authors retain the copyright and assign to the journal the right to the first publication, with the work registered under the attribution, non-commercial and no-derivative license from Creative Commons, which allows third parties to use what has been published as long as they mention the authorship of the work and upon first publication in this journal, the work may not be used for commercial purposes and the publications may not be used to remix, transform or create another work.
b. Authors may enter into additional independent contractual arrangements for the non-exclusive distribution of the version of the article published in this journal (e.g., including it in an institutional repository or publishing it in a book) provided that they clearly indicate that the work was first published in this journal.
c. Authors are permitted and encouraged to publish their work on the Internet (e.g. on institutional or personal pages) before and during the review and publication process, as it may lead to productive exchanges and faster and wider dissemination of published work (see The Effect of Open Access).
















