Fungi associated with dry rot disease of yam (Dioscorea rotundata Poir.) tubers in Cordoba, Colombia
DOI:
https://doi.org/10.15517/am.v32i3.41697Keywords:
diseases, storage, postharvest, Lasiodiplodia theobromae, Fusarium solaniAbstract
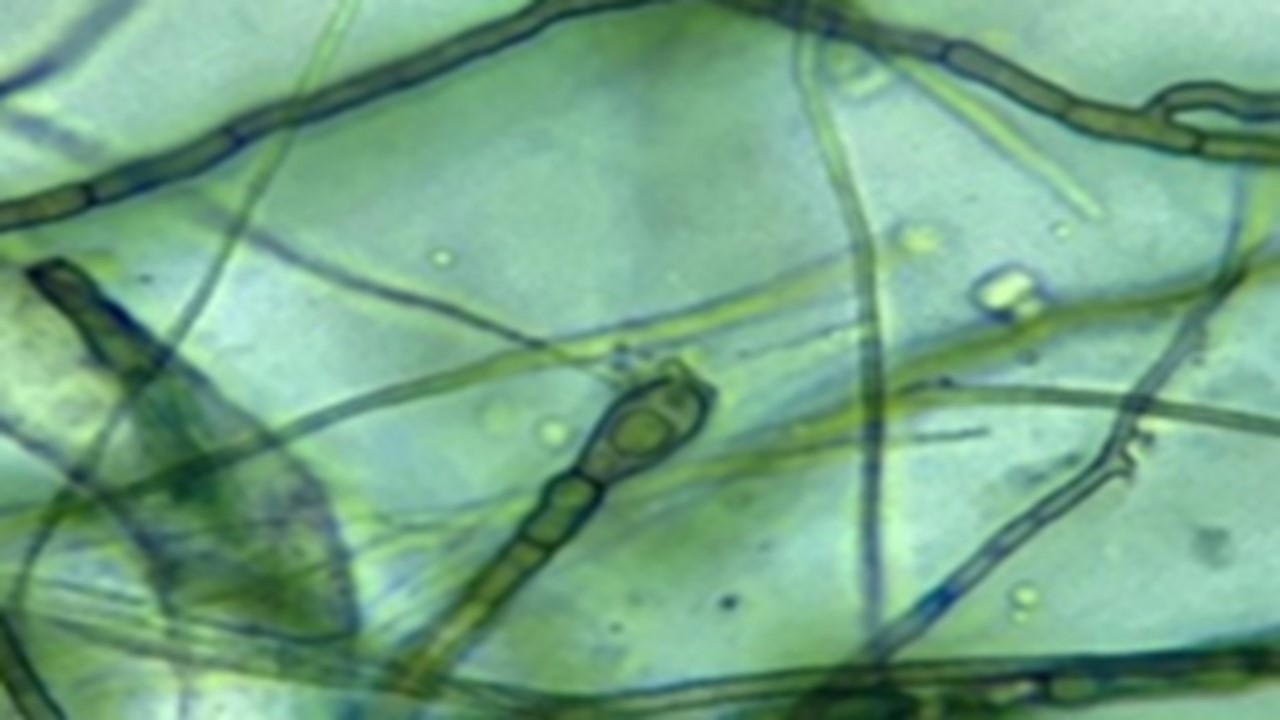
Introduction. Dry rot is a disease that has acquired great importance in Colombia because it causes losses in yam (Dioscorea rotunda Poir.) tubers, both in the field and in postharvest, affecting the economic income and productivity of smallholders. Information on the causal agents of this disease is scarce, so the strategies for managing this problem are unclear. Objective. To identify and characterize fungi associated with the dry rot disease of yam tubers. Materials and methods. This research was carried out between March and October 2019, in the coastal zone of the Córdoba department in the Colombian Caribbean. Yam tubers were collected in storage, due to the fact that farmers keep them as seeds for future harvests. In the laboratory, the associated fungi was isolated from the lesions, characterized, and then identified through the amplification of the ITS (Internal Transcribed Spacer) region of the ribosomal DNA. Results. Twenty-eight fungal growths associated with dry rot of yam tubers were isolated and grouped into eleven morphotypes of which ten species were identified, belonging to nine fungal genera. Conclusion. Six fungi could be associated with the dry rot of yam tubers in the coastal zone of Colombia. These would be: Lasiodiplodia theobromae, Fusarium solani, Rhizopus oryzae, Neocosmospora rubicola, Neoscytalidium dimidiatum, and Curvularia aeria.
Downloads
References
Adeniji, A., Taiga, A., & Ayodele, M. S. (2020). Comparative studies on the susceptibility of three tubers of Dioscorea species to dry rot in Anyigba, Kogi State. International Journal Annals of Science, 8(1), 70–74. https://doi.org/10.21467/ias.8.1.70-74.
Ajayi, A. O., & Olorundare, S. D. (2014). Bacterial and fungal species associated with yam (Dioscorea rotundata) rot at Akungba-Akoko, Ondo State of Nigeria. Applied Science Research Journal, 2(2), 12–28. https://www.researchgate.net/publication/329254575_BACTERIAL_AND_FUNGAL_SPECIES_ASSOCIATED_WITH_YAM_DIOSCOREA_ROTUNDATA_ROT_AT_AKUNGBA-AKOKO_ONDO_STATE_OF_NIGERIA
Ali, S. S., Asman, A., Shao, J., Balidion, J. F., Strem, M. D., Puig, A. S., Meinhardt, L. W., & Bailey, B. A. (2020). Genome and transcriptome analysis of the latent pathogen Lasiodiplodia theobromae, an emerging threat to the cacao industry. Genome, 63(1), 37–52. https://doi.org/10.1139/gen-2019-0112
Amadioha, A. C. (2001). Fungitoxic effects of some leaf extracts against Rhizopus oryzae causing tuber rot of potato. Archive of Phytopathology and Plant Protection, 33(6),499–507. https://doi.org/10.1080/03235400109383372
Arenas, R. (2014). Micología médica ilustrada. Mc Graw Hill.
Arya, R. S., Sheela, M. N., Jeeva, M. L., & Abhilash, P. V. (2019). Identification of Host Plant Resistance to Anthracnose in Greater Yam (Dioscorea alata L.). International Journal of Current Microbiology and Applied Sciences, 8(8), 1690–1696. https://doi.org/10.20546/ijcmas.2019.808.199
Barnet, H. L., & Hunter, B. B. (1998). Illustrated genera of imperfect fungi 4 th. The American Phytopathological Society Press.
Battaglia, E., Benoit, I., Van den Brink, J., Wiebenga, A., Coutinho, P.M., Henrissat, B., & de Vries, R. P. (2011). Carbohydrate-active enzymes from the zygomycete fungus Rhizopus oryzae: a highly specialized approach to carbohydrate degradation depicted at genome level. BMC Genomics, 12(1), Article 38. https://doi.org/10.1186/1471-2164-12-38
Chinchilla, S. G. (2017). Identificación molecular y morfológica de las especies de Fusarium, asociadas al cultivo de la pimienta negra (Piper nigrum L.) en la virgen de Zarapiquí (Tesis Doctoral, no publicada). Universidad de Costa Rica.
Chowdhury, M. K., Jahan, M. S., Akhtar, S., Islam, M. A., Islam, M. A., Sikdar, B., & Hasan, M. F. (2019). Characterization of fungal pathogens causing diseases in bitter gourd and establishment of their eco-friendly control measure. International Journal of Multidisciplinary Research and Development, 6(1), 109-115. https://www.researchgate.net/profile/Md_Hasan151/publication/330423175_Characterization_of_fungal_pathogens_causing_diseases_in_bitter_gourd_and_establishment_of_their_eco-friendly_control_measure/links/5c49434d299bf12be3df2643/ Characterization-of-fungal-pathogens-causing-diseases-in-bitter-gourd-and-establishment-of-their-eco-friendly-control-measure.pdf
Coleman, J. J. (2016). The Fusarium solani species complex: ubiquitous pathogens of agricultural importance. Molecular Plant Pathology, 17(2), 146–158. https://doi.org/10.1111/mpp.12289
Coursey, D. G. (1967). Yam: An account of the nature, origins, cultivation and utilization of the useful member of the Dioscoreaceae. Longmans, Green and Co. Ltd.
Curvelo, G. L., & Rojas, B. A. J. (2010). Revisión preliminar de medios de cultivo empleados en estudios de microorganismos de los phylum Ascomycetes, Deuteromycetes y Oomycetes como agentes causantes de enfermedades en plantas (Tesis de Pregrado, no publicada). Pontificia Universidad Javeriana.
Dania, V. O., Fadina, O. O., Ayodele, M., & Kumar, P. L. (2019). Distribution and virulence of fungal species isolated from yam (Dioscorea spp.) tubers in three agroecological zones of Nigeria. International Journal of Pest Management, 66(3), 1–10. https://doi.org/10.1080/09670874.2019.1629041
De-Pinho, R. S. C., Pozzebon, B. C., Calvano, C. C. A., Vey, R. T., dos Santos Hajar, A., Rodrigues, B. M., & Rodrigues, K. R. R. (2019). Bioprospecção de rizobactérias para o controle in vitro de Pyricularia grisea, tratamento de sementes e promoção de crescimento de plântulas de arroz. Biotemas, 32(3), 23–34. https://doi.org/10.5007/2175-7925.2019v32n3p23
Ezeibekwe, I.O., Umeoka, N., & Izuka, C.M. (2016). Field survey of symptoms and isolation of fungi associated with post-harvest rots of white yam (Dioscorea rotundata Poir.). Journal of Food Processing & Technology, 7(12), 1–4. https://doi.org/10.4172/2157-7110.1000642
Food and Agriculture Organization of the United Nations. (2017). Statistical databases. http://www.fao.org/faostat/en/#data/QC/visualize
Fonseca, J. Y., Castañeda, A. E., Escarraga, J. O., & Cubillos, D. D. (2019). Caracterización de enfermedades fitopatógenas en el cultivo de tomate de árbol (Solanum betaceum) en la finca el reposo en el municipio Facatativá, Cundinamarca. Ciencias Agropecuarias, 5(1), 24–31. https://doi.org/10.36436/24223484.192
Gardes, M., & Bruns, T. D. (1993). ITS primers with enhanced specificity for basidiomicetes- application to the identification of mycorrhizae and rusts. Molecular Ecology, 2(2), 113–118. https://doi.org/10.1111/j.1365-294X.1993.tb00005.x
Garzón, N. (2013). Caracterización e identificación molecular de hongos de suelo aislados de los páramos de Guazca y Cruz verde, Cundinamarca-Colombia (Tesis de Pregrado, no publicada). Pontificia Universidad Javeriana.
Gwa, V. I., & Ekefan, E. J. (2017). Fungal Organisms Isolated from Rotted White Yam (Dioscorea rotundata) Tubers and Antagonistic Potential of Trichoderma harzianum against Colletotrichum Species. Agricultural Research & Technology: Open Access Journal, 10(3), Article 555787. https://doi.org/10.19080/ARTOAJ.2017.10.555787
Gwa, V. I., & Ekefan, E. J. (2018). Fungicidal effect of some plant extracts against tuber dry rot of white yam (Dioscorea rotundata) Caused by Aspergillus Niger. International Journal of Horticulture & Agriculture, 3(3), Article 123. https://www.researchgate.net/publication/330365381_Fungicidal_Effect_of_Some_Plant_Extracts_against_Tuber_Dry_Rot_of_White_Yam_Dioscorea_Rotundata_Poir_Caused_by_Aspergillus_niger
Gwa, V. I., & Richard, I. B. (2018). Susceptibility of white yam (Dioscorea rotundata) tuber to rot fungi and control with extracts of Zingiber officinale Rosc. Azadirachta indica A. Juss. and Piper guineense Schumach. Journal of Plant Pathology & Microbiology, 9(9), Article 452. https://doi.org/10.4172/2157-7471.1000452
Hernández-Trejo, A., Estrada-Drouaillet, B., López-Santillán, J. A., Rios-Velasco, C., Varela-Fuentes, S. A., Rodríguez-Herrera, R., & Osario-Hernández, E. (2019). In vitro evaluation of native entomopathogenic fungi and neem (Azadiractha indica) extracts on Spodoptera frugiperda. Phyton, International Journal of Experimental Botany, 88(1), 47–54. http://doi.org/10.32604/phyton.2019.04611
Jovičić-Petrović, J., Jeremić, S., Vučković, I. M., Vojnović, S., Bulajic, A., Raicevic, V., & Nikodinović-Runić, J. (2016). Aspergillus piperis A/5 from plum-distilling waste compost produces a complex of antifungal metabolites active against the phytopathogen Pythium aphanidermatum. Archives of Biological Sciences, 68(2), 279–289. https://doi.org/10.2298/ABS150602016J
Kim, J. H., Kim, D.Y., Park, H., Cho, J. H., & Eom, A. H. (2017). Neocosmospora rubicola, an unrecorded endophytic fungus isolated from roots of Glycyrrhiza uralensis in Korea. The Korean Journal of Mycology, 45(1), 63-67. https://doi.org/10.4489/KJM.20170007
Lin, C. H., Chen, Y. X., Liu, W. B., Wu, W. Q., Miao, W. G., & Zheng, F. C. (2017). First report of Dioscorea esculenta dieback caused by Neoscytalidium dimidiatum in China. Plant Disease, 101(7), Article 1320. https://doi.org/10.1094/PDIS-02-17-0167-PDN
Lombard, L., Van der Merwe, N. A., Groenewald, J. Z., & Crous, P. W. (2015). Generic concepts in Nectriaceae. Studies in Mycology, 80, 189–245. https://doi.org/ 10.1016/j.simyco.2014.12.002
Londoño-Hernández, L., Ramírez-Toro, C., Ruiz, H. A., Ascacio-Valdés, J. A., Aguilar-Gonzalez, M. A., Rodríguez-Herrera, R., & Aguilar, C. N. (2017). Rhizopus oryzae–Ancient microbial resource with importance in modern food industry. International Journal of Food Microbiology, 257, 110–127. https://doi.org/10.1016/j.ijfoodmicro.2017.06.012.
Ministerio de Agricultura y Desarrollo Rural. (2018). Indicadores e instrumentos del sector productivo del ñame en Colombia. https://sioc.minagricultura.gov.co/Yuca/Documentos/2018-09-30%20Cifras%20Sectoriales.pdf
Mello, J. F., Brito, A. C. Q., Motta, C. M. S., Vieira, J. C. B., Michereff, S. J., & Machado, A. R. (2019). First report of Neoscytalidium dimidiatum causing root rot in sweet potato in Brazil. Plant Disease, 103(2), Article 373. https://doi.org/10.1094/PDIS-07-18-1242-PDN
Montiel, M. F., Isla, L. H., & Romero, L. M. (2016). Microorganismos fitopatógenos y asociados a tubérculos de cultivares de ñame (Dioscorea sp.). Agricultura Tropical, 2(1), 61–68. http://ojs.inivit.cu/index.php?journal=inivit&page=article&op=view&path%5B%5D=46&path%5B%5D=AT02012016_7MP-011.pdf
Munirah, M. S., Azmi, A. R., Yong, S. Y. C., & Nur Ain Izzati, M. Z. (2017). Characterizationof Lasiodiplodia theobromae and L. pseudotheobromae causing fruit rot on rot on pre-harvest mango in Malaysia. Plant Pathology & Quarantine, 7(2), 202–213. https://doi.org/10.5943/ppq/7/2/14
Nayab, M., & Akhtar, N. (2016). New report of Cycas revoluta leaf necrosis by Phoma herbarum from Pakistan. Journal of Plant Diseases and Protection, 123(4), 193–196. https://doi.org/10.1007/s41348-016-0026-z
O’Donnell, K., & Cigelnik, E. (1997). Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the FungusFusariumAre nonorthologous. Molecular Phylogenetics and Evolution, 7(1), 103–116. https://doi.org/10.1006/mpev.1996.0376
O’Donnell, K., Nirenberg, H. I., Aoki, T., & Cigelnik, E. (2000). A multigene phylogeny of the Gibberella fujikuroi species complex: detection of additional phylogenetically distinct species. Mycoscience, 41(1), 61–78. https://doi.org/10.1007/BF02464387
Osorio-Gutiérrez, L. A., & Castaño-Zapata, M. (2011). Caracterización del agente causante de la pudrición de raíces de la arveja (Pisum sativum linneo), enfermedad endémica en el municipio de Manizales Caldas (Colombia). Agronomía, 19(2), 33–43. http://agronomia.ucaldas.edu.co/downloads/Agronomia19(2)_4.pdf
Pacasa-Quisbert, F., Loza-Murguia, M.G., Bonifacio-Flores, A., Vino-Nina, L., & Serrano-Canaviri, T. (2017). Comunidad de hongos filamentosos en suelos del Agroecosistema de K’iphak’iphani, Comunidad Choquenaira-Viacha. Journal of the Selva Andina Research Sociaty, 8(1), 2–25. http://www.scielo.org.bo/pdf/jsars/v8n1/v8n1_a02.pdf
Peixinho, G. D. S., Ribeiro, V. G., Amorim, E. P. D. R., & Morais, A. C. D. M. (2019). Ação do óleo essencial de Citronela (Cymbopogon nardus L) sobre o patógeno Lasiodiplodia theobromae em cachos de videira cv. Itália. Summa Phytopathologica, 45(4), 428–431.
Purseglove, J. W. (1972). Tropical crops-monocotyledons London. Longmans Green and Co. Ltd.
Rojas, L., Cajiao, A., & Rivera, K. (2019). Estudio y análisis presuntivo de la fertilidad microbiana del suelo de un cultivo de maíz (Zea mays) en Pamplona, Norte de Santander. Bistua: Revista de la Facultad de Ciencias Básicas, 16(1), 3–12. http://revistas.unipamplona.edu.co/ojs_viceinves/index.php/BISTUA/article/view/3186/1697
Rosado, I. Y. (2016). Evaluación de fungicidas orgánicos y convencionales para el control de enfermedades foliares en ñame (Dioscorea alata L.) (Tesis Maestría, no publicada). Universidad de Puerto Rico.
Salcedo-Morales, G., Trejo-Espino, J. L., Martínez-Bonfil, B. P., Cruz-Sosa, F., & Trejo-Tapia, G. (2017). Formación de raíces e inducción de haustorios de Castilleja tenuiflora Benth. con catequina y peróxido de hidrógeno. Polibotánica, 44, 147–157. https://doi.org/10.18387/polibotanica.44.11
Sánchez-López, D. B., & Pérez-Pazos, J. V. (2018). Caracterización y evaluación de PGPRs sobre el crecimiento de plántulas de Dioscorea rotundata in vitro. Agronomía Costarricense, 42(2), 75–91. https://doi.org/10.15517/rac.v42i2.33780
Sandoval-Denis, M., & Crous, P. W. (2018). Removing chaos from confusion: assigning names to common human and animal pathogens in Neocosmospora. Persoonia, 41, 109–129. https://doi.org/10.3767/persoonia.2018.41.06
Sreevidya, M., & Gopalakrishnan, S. (2016). Penicillium citrinum VFI-51 as biocontrol agent to control charcoal rot of sorghum (Sorghum bicolor (L.). African Journal of Microbiology Research, 10(19), 669–674. https://doi.org/10.5897/AJMR2015.7831
Shiriki, D., Ubwa, S. T., y Shambe, T. (2015). Isolation of nine microorganisms from rotten Dioscorea rotundata (white yam) and antimicrobial sensitivity test with five plant extracts. Food and Nutrition Sciences, 6(10), 825–835. https://doi.org/10.4236/fns.2015.610086
Villa-Martínez, A., Pérez-Leal, R., Morales-Morales, H. A., Basurto-Sotelo, M., Soto-Parra, J. M., & Martínez-Escudero, E. (2015). Situación actual en el control de Fusarium sp. y evaluación de la actividad antifúngica de extractos vegetales. Acta Agronómica, 64(2), 194–205. https://doi.org/10.15446/acag.v64n2.43358
Voigt, K., Cigelnik, E., & O’donnell, K. (1999). Phylogeny and PCR identification of clinically important Zygomycetes based on nuclear ribosomal-DNA sequence data. Journal of Clinical Microbiology, 37(12), 3957–3964. https://doi.org/10.1128/JCM.37.12.3957-3964.1999
Waqas, M., Khan, A. L., Hamayun, M., Shahzad, R., Kang, S. M., Kim, J. G., & Lee, I. J. (2015). Endophytic fungi promote plant growth and mitigate the adverse effects of stem rot: An example of Penicillium citrinum and Aspergillus terreus. Journal of plant interactions, 10(1), 280–287. https://doi.org/10.1080/17429145.2015.1079743
Zheng, F., Xu, G., Zheng, F. Q., Ding, X. F., & Xie, C. P. (2018). Neocosmospora rubicola causing stem rot of Pitaya (Hylocereus costaricensis) in China. Plant Disease, 102(12), 2653-2653. https://doi.org/10.1094/PDIS-09-17-1469-PDN
Downloads
Additional Files
Published
How to Cite
Issue
Section
License
1. Proposed policy for open access journals
Authors who publish in this journal accept the following conditions:
a. Authors retain the copyright and assign to the journal the right to the first publication, with the work registered under the attribution, non-commercial and no-derivative license from Creative Commons, which allows third parties to use what has been published as long as they mention the authorship of the work and upon first publication in this journal, the work may not be used for commercial purposes and the publications may not be used to remix, transform or create another work.
b. Authors may enter into additional independent contractual arrangements for the non-exclusive distribution of the version of the article published in this journal (e.g., including it in an institutional repository or publishing it in a book) provided that they clearly indicate that the work was first published in this journal.
c. Authors are permitted and encouraged to publish their work on the Internet (e.g. on institutional or personal pages) before and during the review and publication process, as it may lead to productive exchanges and faster and wider dissemination of published work (see The Effect of Open Access).




















