Morphological, morphometric and molecular identification of Meloidogyne exigua (Göeldi 1887) in coffee (Coffea arabica)
DOI:
https://doi.org/10.15517/am.v31i3.38733Keywords:
Coffea arabica, root-knot nematode, identification, molecular analysisAbstract
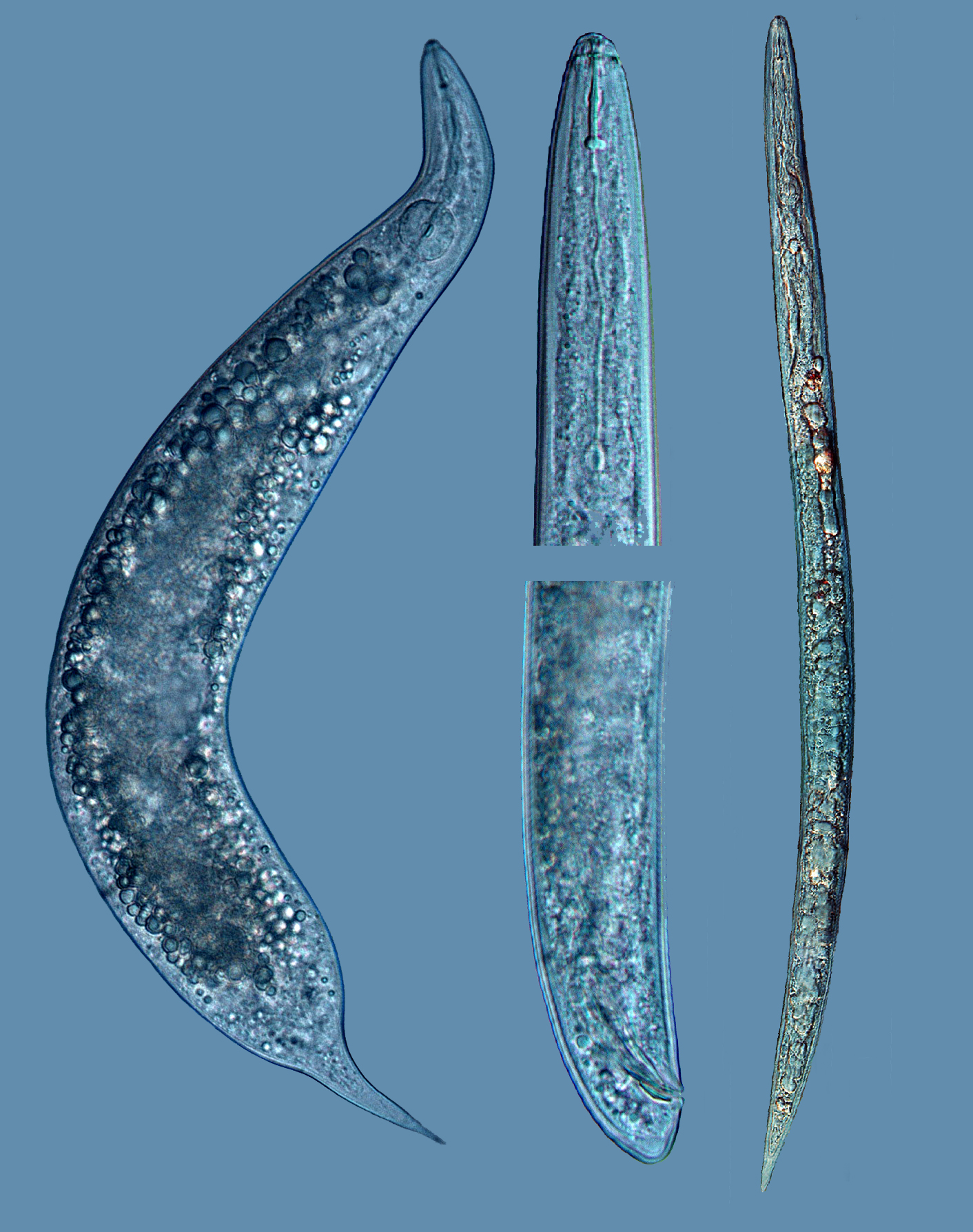
Introduction. The genus Meloidogyne contains many species and their correct diagnosis of these must be based on the use of multiple tools, which in diagnostic laboratories allow a correct and reliable evaluation, specially for quarantine nematodes. The proper identification of a phytonematode is important not only to establish an appropriate control strategy, but also to avoid its spread to other areas where it could cause damage that would result in economic losses. Objective. To perform taxonomic and molecular studies to confirm the Meloidogyne species, associated to a coffee plantation located in Barva, Heredia. Materials and methods. Soil and root samples were collected in a coffee plantation located in Barva, Heredia, Costa Rica, which were processed in the Nematology laboratory of the School of Agricultural Sciences at the Universidad Nacional (UNA), using the centrifugation-flotation method in sugary solution and molecular techniques. Results. On average, 30 J2 100 cc-1 were counted in soil and 1000 J2 10 g-1 of roots of the root-knot nematode. Taxonomic and molecular techniques allowed to identify that the females and juveniles extracted from coffee roots corresponded to Meloidogyne exigua. Analysis of the sequences generated by PCR and the use of the DraI enzyme confirmed the identity of the species with 99 % similarity when compared to sequences from Costa Rica and Nicaragua, reported in the Genbank. Conclusion. Using different morphometric and molecular techniques, M. exigua associated with a coffee plantation was identified.
Downloads
Downloads
Additional Files
Published
How to Cite
Issue
Section
License
1. Proposed policy for open access journals
Authors who publish in this journal accept the following conditions:
a. Authors retain the copyright and assign to the journal the right to the first publication, with the work registered under the attribution, non-commercial and no-derivative license from Creative Commons, which allows third parties to use what has been published as long as they mention the authorship of the work and upon first publication in this journal, the work may not be used for commercial purposes and the publications may not be used to remix, transform or create another work.
b. Authors may enter into additional independent contractual arrangements for the non-exclusive distribution of the version of the article published in this journal (e.g., including it in an institutional repository or publishing it in a book) provided that they clearly indicate that the work was first published in this journal.
c. Authors are permitted and encouraged to publish their work on the Internet (e.g. on institutional or personal pages) before and during the review and publication process, as it may lead to productive exchanges and faster and wider dissemination of published work (see The Effect of Open Access).
















